Flekklavfamilien
Arthoniaceae











Flekklavfamilien er en artsrik familie som omfatter både skorpelav og lavboende sopper, ofte kalt lavparasitter, samt en del saprofytter som lever på bark. Totalt er det på verdensbasis kjent ca 850 arter, og de finnes fra tropiske til arktisk-alpine områder. Fra Norge er det så langt rapportert 76 arter i familien. Blant skorpelavene i flekklavfamilien er det en rekke sjeldne arter som er karakteristiske for fuktige skoger med lang kontinuitet. Spesielt i de sørvestlige regnskogene i Norge er det mange flekklav-arter. Kun lavdannende og barklevende sopp i flekklavfamilien omhandles her.
Beskrivelse
Thallus
Tallus hos flekklavene er skorpeforma og ofte dårlig utvikla. I de fleste tilfellene er tallus svært enkelt oppbygd og det er som regel ikke mulig å se noen differensiering i barklag, marg og algesjikt som hos bladlavene. Hos noen arter kan tallus være forholdsvis tykt på utsida av substratet, mens det hos andre arter er utydelig eller helt innleira i substratet. I slike tilfeller kan en bare se en svak farging av substratet. Kanten av tallus er enten utydelig avgrensa eller avgrensa av en tynn brunsvart linje. Algekomponenten hører enten til grønnalgefamilien Trentepohliaceae som ofte danner flercella kolonier med guloransje pigmenter eller det kan være encella runde grønnalger som gjerne kalles chlorococcoide alger.
Fruktlegemer
Fruktlegemene hos flekklavene er av typen apothecier. De kan være nokså varierende i utseende, men framtrer ofte som mer eller mindre runde flekker eller de kan være avlange og smale, til dels forgreina eller stjerneforma. De kan være flate eller svakt konvekse og mer eller mindre innsenka i tallus. I noen tilfeller kan de være delvis opphøyet og tilsynelatende festet til tallusoverflata. Fargen varierer fra nyanser av brunoransje, rødbrunt og brunsvart til svart. Hos noen arter er overflata av fruktlegemene dekket av et fint lag av krystaller (pruina) som kan være hvitt, oransje eller rødt.
Karakteristisk for de fleste artene i flekklavfamilien er at fruktlegemene mangler en markert, velavgrensa kant. Det finnes imidlertid noen unntak som for eksempel svartflekklav Arthonia atra, kalkflekklav Arthonia calcarea, kantflekklav Arthonia excipienda og granittflekklav Arthonia granitophila. Hos disse er det en hard, brunsvart kant.
Det sporeproduserende vevet (hymeniet) hos flekklavene består av forgreina eller nettforma hyfer (parafysoider) som er omgitt av en fargeløs eller svakt pigmentert gelatinøs matrix. Spissene på parafysoidene er ofte utvida og har pigmenter og hos de fleste artene danner de et tydelig sjikt (epithecium) på toppen av hymeniet. Hos noen arter står endene på parafysoidene loddrett mens hos andre bøyer de av og ligger horisontalt på overflata av hymeniet. Pigmentene opptrer enten som en diffus oppløsning eller som bittesmå gryn. Sjiktet på undersiden av hymeniet (hypothecium) er enten fargeløst, mørkt brunt eller rødoransje.
Sporesekkene (asci) er av to typer, enten Arthonia-typen eller Arthothelium-typen. Den indre veggen i sporesekken er fortykket i enden (tholus) og er vanligvis gjennomtrengt av en triangulær til rektangular kanal (ocular chamber). Alle delene av sporesekkene forblir fargeløse ved tilsetning av jod (I) og kaliumjodid-løsning (KI) bortsett fra en ringstruktur i tholus som farges blå hos de fleste artene. Antall sporer i hver sporesekk er normalt 8, men kan variere fra 1 til 8 avhengig av arten.
Sporene hos flekklavene er 2- til mangecellet. De kan enten ha bare tverrvegger eller de kan i tillegg også ha langsgående vegger. I det siste tilfellet snakker vi om muriforme sporer. I begge tilfeller er det hos mange arter en tydelig forstørra endecelle. Sporene er enten fargeløse hele tiden eller de kan bli brunpigmenterte med alderen.
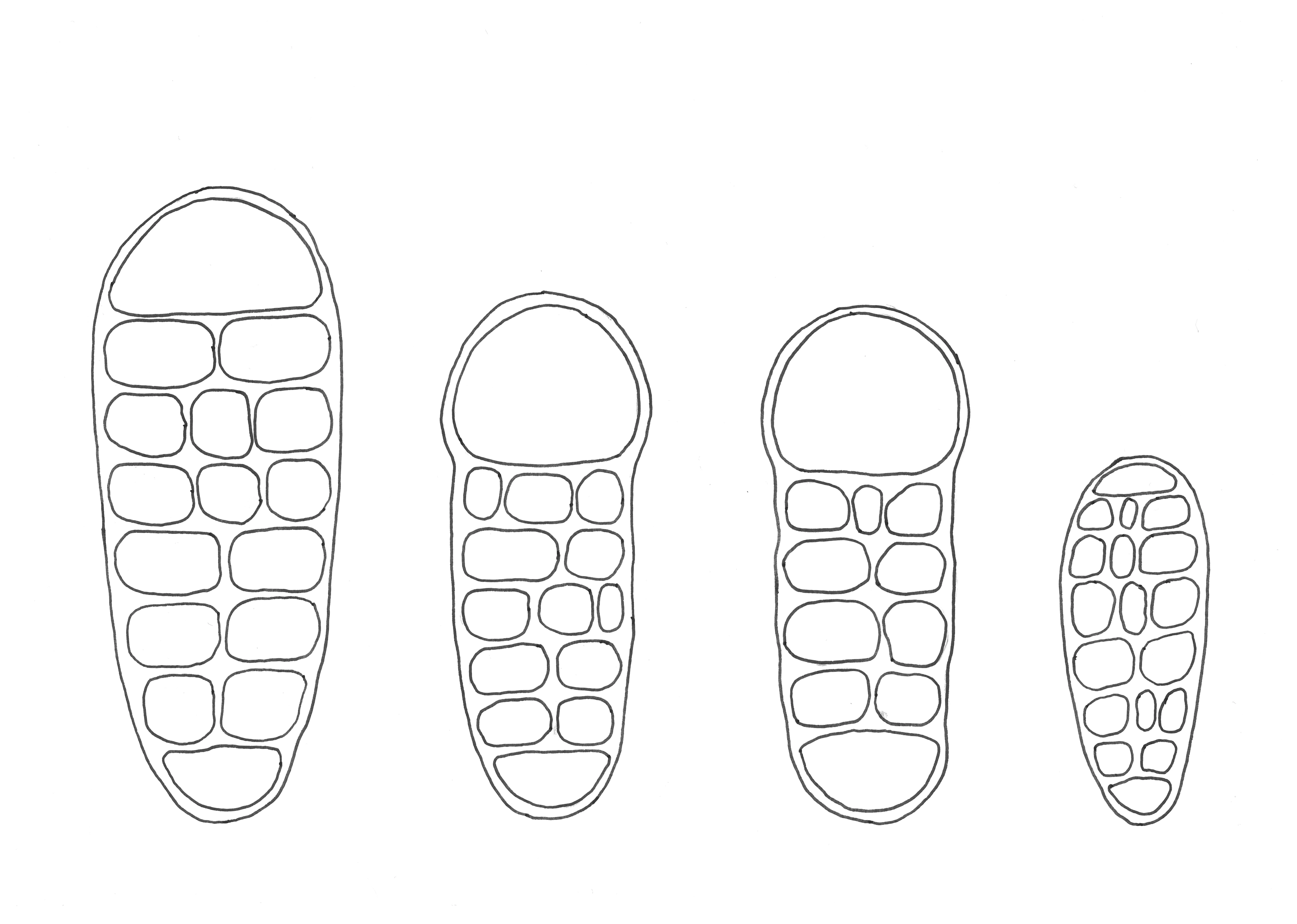
Figur 1. Sporetegninger, fra venstre mot høyre: Trønderflekklav Arthothelium norvegicum, nettsporeflekklav Arthothelium dictyosorum, hasselflekklav Arthothelium macounii og jaguarflekklav Arthothelium ruanum. Sporen til nettsporeflekklav er 30 µm lang.
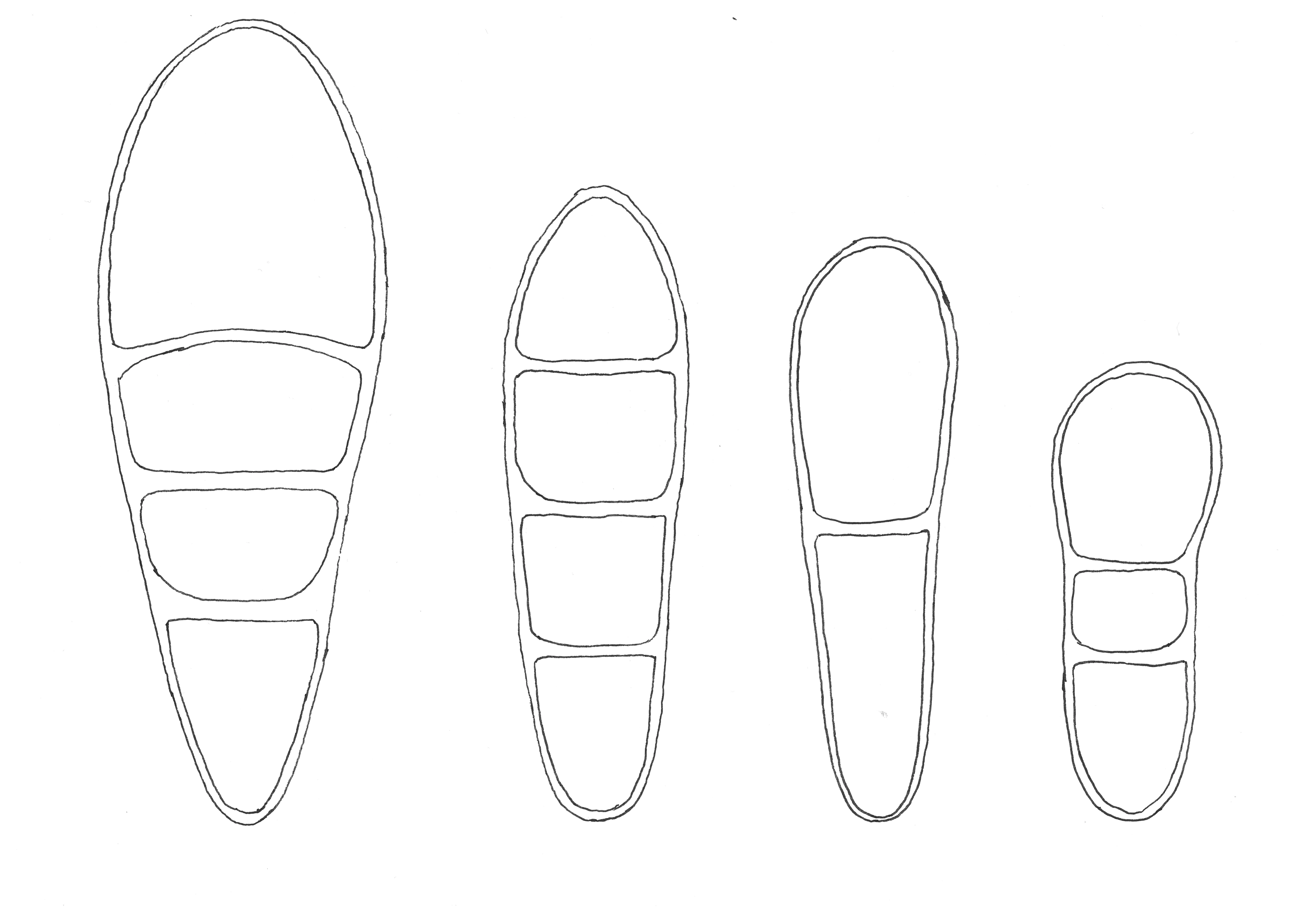
Figur 2. Sporetegninger, fra venstre mot høyre: Stjerneflekklav Arthonia stellaris, stråleflekklav Arthonia radiata, vierflekklav Arthonia apatetica og honningflekklav Arthonia helvola. Sporen til honningflekklav er 10 µm lang.
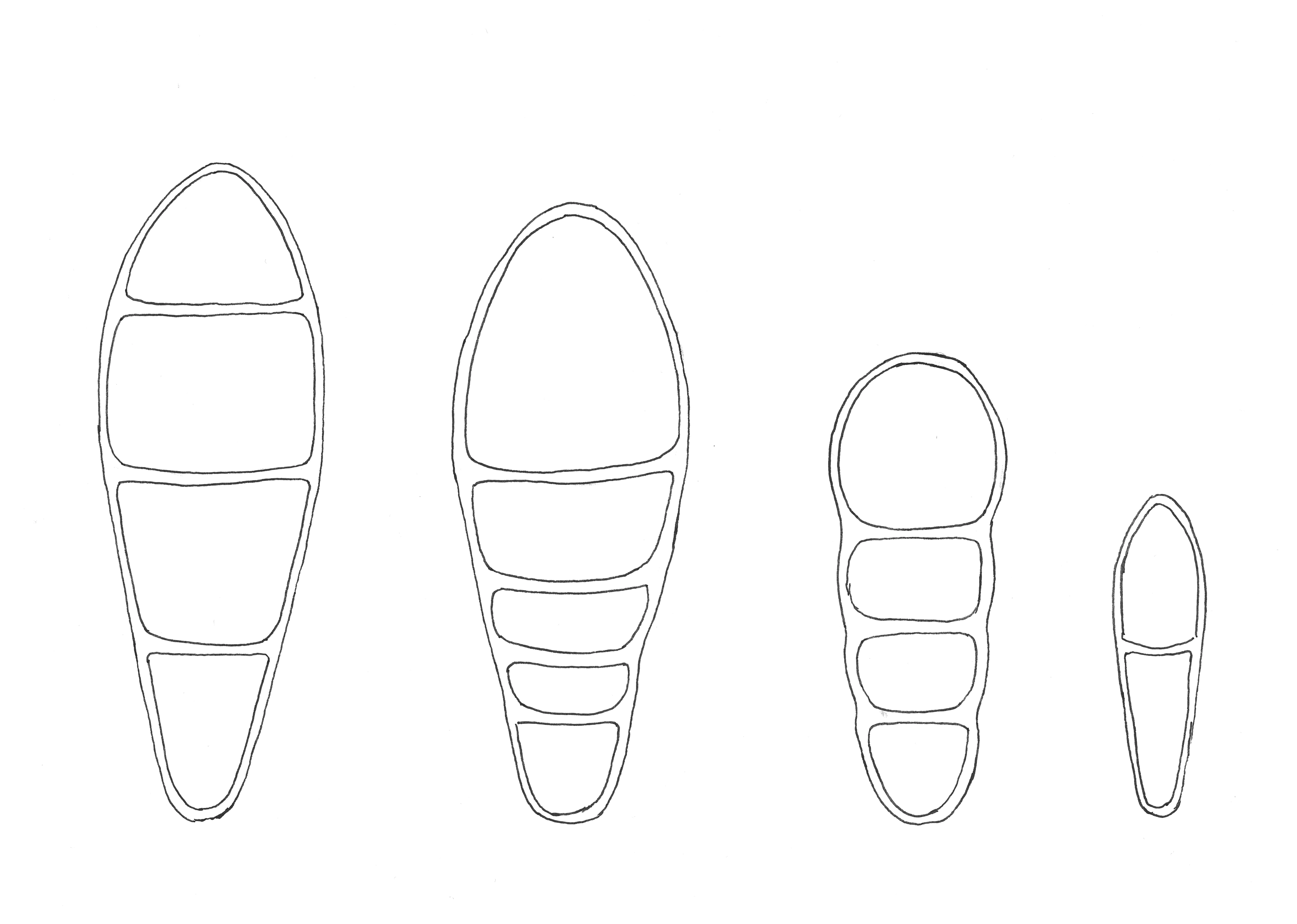
Figur 3. Sporetegninger, fra venstre mot høyre: Kvistflekklav Naevia punctiformis, praktflekklav Coniocarpon fallax, eremittflekklav Reichlingia anombrophila og moseflekklav Bryostigma muscigenum. Sporen til moseflekklav er 10 µm lang.
Ukjønna formering
Ukjønna sporer som dannes ved vanlig celledeling (mitose) kalles konidier. Hos norske flekklaver dannes konidier inne i små flaskeforma beholdere (pyknidier). Konidiene er stavforma, ellipseforma eller trådforma. Andre typer av konidie-produserende organ forekommer hos flere arter av flekklaver andre steder i verden.
Kjemi
Mange av de norske flekklavene mangler lavsyrer, såkalte sekundære metabolitter, i tallus og fruktlegemer. Hos noen arter derimot finnes en ganske stor variasjon av anthraquinon-pigmenter og depsider. Funksjonen til lavsyrene i lavsymbiosen er ikke alltid fullgodt klarlagt, men det er dokumentert at noen beskytter mot beiting fra for eksempel snegler og insekter. Andre lavsyrer beskytter mot bakterier og virus mens andre igjen beskytter mot skadelig UV-lys. Det er også vist at enkelte lavsyrer beskytter mot overmetning med vann slik at utvekslinga av karbondioksid og oksygen i fotosyntesen kan fungere optimalt.
Lavsyrene kan påvises ved hjelp av tynnsjiktkromatografi (TLC) eller enkle fargereaksjoner. Fargereaksjoner på tallus og fruktlegemer kan gjennomføres med noen enkle reagenser og en noterer hvilke fargeendringer som oppstår. De vanligste reagensene til dette formålet er:
- K: 10 % løsning av kaliumhydroksid i vann
- C: klorholdig blekemiddel – vanlig klorin
- KC: reagens K umiddelbart etterfulgt av reagens C
- PD: en mettet løsning av parafenylendiamin i alkohol
- I: Lugols jodløsning
- KI: Lugols jodløsning etter forbehandling med reagens K
Testing for amyloide reaksjoner (I+ blå) eller hemiamyloide reaksjoner (I+ rød, KI+ blå) i fruktlegemenes gelatinøse matrix er spesielt viktig for den systematiske klassifiseringa hos flekklavene.
Økologi
De fleste flekklavene i Norge opptrer som epifytter i skog i lavlandet, særlig langs kysten. Store, gamle trær i parker og alléer er også et viktig habitat for flekklaver. De fleste artene vokser på bark av løvtrær, særlig edelløvtrær, men noen forekommer også, eller bare, på bartrær som gran og furu. I lavsamfunnene opptrer de fleste flekklavene som pionerarter på unge stammer eller kvister med glatt bark, men noen opptrer på gamle trær i skog med lang kontinuitet og kan derfor signalisere skog med høy bevaringsverdi. Noen flekklaver er vanlige, vidt utbredte arter, mens andre har en klart sørlig eller sørvestlig utbredelse i Norge. Flere flekklaver er eksklusive regnskogsarter enten i boreonemorale eller boreale regnskoger langs den norske vestkysten. Det er kun noen få steinboende flekklaver og disse vokser ofte godt beskytta på overhengende bergvegger i skoglandskapet. Enkelte arter kan sågar vokse på menneskeskapte substrater som betong, takstein og murvegger eller på mer eller mindre kalkrikt berg i fjæresonen.
De fleste lavboende soppene i flekklavfamilien tilhører slekta Arthonia. I motsetning til de lavdannende artene i familien, er denne gruppen også vanlig og vidt utbredt i alpine habitater. De fleste artene er sterkt bundet til en bestemt vert eller til noen få nært beslekta lavarter. Lavboende arter i flekklavfamilien er i Norge kjent fra en rekke ulike lavarter innenfor både bladlav, busklav og skorpelav.
Systematikk
Flekklavfamilien Arthoniaceae er den største familien blant sju aksepterte familier innenfor flekklavordenen Arthoniales. Sammen med orden Lichenostigmatales, som kun omfatter ikke-lavdannende arter, hører den til soppklassen Arthoniomycetes. Avgrensinga av flekklavfamilien med inndeling i naturlige grupper basert på slektskap er fortsatt under utredning og er derfor ikke endelig fastlagt. Mange slekter er nylig skilt ut fra de store slektene Arthonia og Arthothelium basert på en kombinasjon av morfologiske karakterer og molekylære studier. Eksempler på slike slekter er Bryostigma, Coniocarpon, Diarthonis, Felipes, Inoderma, Leprantha, Naevia, Pachnolepia og Reichlingia. Noen av disse slektene er nylig beskrevet mens andre er gamle slektsnavn som er blitt gjenopptatt.
Selv etter disse omfattende endringene er slektene Arthonia og Arthothelium i Norge å betrakte som heterogene. Slektene er nokså kunstig avgrenset basert på sporetype, sporer med bare tverrvegger hos Arthonia og muriforme sporer hos Arthothelium. Derfor må vi forvente ytterligere endringer av klassifiseringa innenfor flekklavfamilien som også berører norske arter.
Mange av artene som i denne oversikten er plassert i Arthonia har sterke morfologiske likheter med slekta Bryostigma. Fylogenetiske studier viser at de er nært beslektet med Bryostigma muscigenum som er typearten for slekta. De fleste lavboende artene i slekta Arthonia som er kjent fra Norge, hører til denne gruppen. På dette tidspunktet har vi valgt å beholde disse artene i slekta Arthonia i påvente av en skikkelig revisjon av hele denne evolusjonære linjen.
Bestemmelsesnøkler
Mange flekklavarter kan bestemmes i felt basert på ytre morfologi i kombinasjon med det bestemte habitatet arten forekommer i. I mange tilfeller er det imidlertid nødvendig med mikroskopering av indre strukturer i fruktlegemene for å få en sikker bestemmelse. For enkelte arter må også lavsyrene bestemmes.
Følgende bestemmelsesnøkler kan benyttes for å bestemme flekklaver i Norge, men en må være oppmerksom på at ingen av nøklene hver for seg inkluderer alle artene som er kjent fra Norge:
- Arthoniales: Arthoniaceae. Revisions of British and Irish Lichens 1 (Cannon m.fl. 2020)
- Svenska Skorplavar (Foucard 2001)
- Die Flechten Deutschlands, Band 1 (Wirth m.fl. 2013)
Litteratur
Cannon P, Ertz D, Frisch A, Aptroot A, Chambers S, Coppins BJ, Sanderson N, Simkin J og Wolseley P (2020). Arthoniales: Arthoniaceae. Revisions of British and Irish Lichens 1: 1–48. [https://britishlichensociety.org.uk/identification/lgbi3]
Foucard T (2001). Svenska Skorplavar. Interpublishing, Stockholm, 392 pp.
Frisch A, Ohmura Y, Ertz D og Thor G (2015). Inoderma and related genera in Arthoniaceae with elevated white pruinose pycnidia or sporodochia. Lichenologist 47: 233–256.
Frisch A, Thor G, Ertz D og Grube M (2014). The Arthonialean challenge: restructuring Arthoniaceae. Taxon 63: 727–744.
Frisch A, Thor G og Sheil D (2014). Four new Arthoniomycetes from Bwindi Impenetrable National Park, Uganda. Nova Hedwigia 98: 295–312.
Stornes Moen V (2019). Molecular systematics and species delimitation in Coniocarpon and Arthonia punctiformis s.lat. in Norway. Master’s thesis, NTNU, Faculty of Natural Sciences, Department of Natural History, 61 pp.
Sundin R (1999). Phylogenetic and taxonomic studies within Arthonia Ach. (Ascomycetes, Arthoniales). Doctoral dissertation, Department of Bot., Stockholm University.
Sundin R og Tehler A (1998). Phylogenetic studies of the genus Arthonia. Lichenologist 30: 381–413.
Thiyagaraja V, Lücking R, Ertz D, Wanasinghe DN, Karunarathna SC, Camporesi E og Hyde KD (2020). Evolution of non-lichenized, saprotrophic species of Arthonia (Ascomycota, Arthoniales) and resurrection of Naevia, with notes on Mycoporum. Fungal Diversity 102: 205–224.
Wirth V, Hauck M and Schultz M (2013). Die Flechten Deutschlands, Band 1. Ulmer, Stuttgart, 672 pp.