
Moser
Anthocerotophyta, Marchantiophyta og Bryophyta
Norge har en av de mest artsrike mosefloraene i Europa. Dette skyldes først og fremst den store variasjonen i klima, topografi og substrat, som gir opphav til stor naturtypevariasjon. Moser vokser på de fleste underlag, som jord, stein, og ved. De forekommer i de aller fleste landhabitater i Norge, fra lavland til høgfjell, inkludert menneskeskapte habitater. Fra Fastlands-Norge kjenner vi ca. 1140 mosearter. Totalt 322 arter, som utgjør om lag 28 % av den norske mosefloraen, er oppført på Rødlista.
- Innhold
- Kunnskapsgrunnlaget
- Avgrensninger og definisjoner
- Antall rødlistearter
- Årsak til rødlisting
- Endringer fra 2015 til 2021
- Ekspertkomiteen
- Takk
- Referanser
- Siden siteres som:
Navnet moser (bryofytter) er en tradisjonsbetegnelse på tre evolusjonære linjer av landplanter uten karstrenger (indre ledningsvev). De tre hovedgruppene av moser skilles ut på rekkenivå som bladmoser (Bryophyta), levermoser (Marchantiophyta) og nålkapselmoser (Anthocerotophyta). Det har de siste tiårene vært diskutert frem og tilbake om disse tre linjene utgjør ei helhetlig gruppe med felles opphav eller ikke. Nyere analyser gir støtte for at mosene er hverandres nærmeste slektninger med felles stamform (Puttick mfl. 2018).
Moser er grønne med oftest flerårige planteskudd som vanligvis har blader. Disse planteskuddene kalles gametofytter og har et enkelt sett med kromosomer. Det er vanligvis de grønne gametofyttene vi legger merke til og omtaler som moser. Kjønnet reproduksjon hos moser skjer når gametofyttene produserer kjønnsceller. De hannlige kjønnscellene hos moser er, i motsetning til pollen, avhengig av vann for å bevege seg. Spredningsavstanden er derfor begrenset. Ved befruktning dannes en kortlivet sporofytt med dobbelt sett av kromosomer på morplanten. Det er sporofytten som produserer sporer og disse blir dannet i sporehuset på sporofytten. Antall sporer i hvert sporehus varierer fra art til art, og generelt er det slik at arter enten produserer mange små eller få store sporer. Sporene er tilpasset vindspredning, med få unntak, og sporestørrelsen er ofte avgjørende for hvor langt de kan spre seg. Arter med store sporer har gjerne begrenset spredningsevne og sporene er avhengig av at det finnes egnede voksesteder i nærheten for at de skal kunne etablere seg. Dersom en spore havner på et egnet substrat, kan den spire og gi opphav til en ny gametofytt (moseskudd).
Mosene har også ukjønnet formering – enten gjennom gjentatt forgreining av de grønne moseskuddene, dannelse av skuddfragmenter eller ved produksjon av ulike former for grokorn som kan vokse opp til nye moseplanter. Disse vegetative enhetene er ofte større og tyngre enn sporer, men kan spre seg effektivt med andre vektorer enn vind, som for eksempel med fugl og pattedyr. Vegetative skudd er også mindre kravfulle til substratet ved etablering enn sporer. Moseskudd forgreiner seg og vil over tid dele seg i separate moseplanter. Det er derfor vanlig å anta at moseskudd som står tett, tilhører den samme klonen.
Mosene tar opp vann og næringssalter gjennom hele overflaten, de har ikke ekte røtter og mangler eller har dårlig utviklet ledningsvev (karstrenger) sammenlignet med karplantene. De fleste mosene tåler kortere eller lengre perioder med uttørking av skuddene; da går de inn i en dvaletilstand. Når de igjen får tilgang på vann, gjenopptar de livsfunksjonene. Dette gjør at mange moser kan vokse på svært tørre steder, for eksempel på sørvendte bergvegger. En motsatt strategi er å unngå uttørking, en tilpasning som er vanligere hos moser som vokser i våte habitater.
Noen få arter vokser på møkk, smågnagerkadavre og andre dyrerester (møkkmoser og lemenmoser). En del arter forekommer også i ferskvann. Moser dominerer i flere naturtyper, for eksempel på myr og i skogbunnen. I disse økosystemene har mosene viktige økologiske funksjoner. Relativt sett er mosene viktigere i åpne naturtyper uten stor karplantedominans, som i myr, fjellet og arktiske områder, både med tanke på mangfold av arter og produksjon av biomasse (Hassel mfl. 2012, Longton 1988). Omkring 40 % av Norges moser, og rundt 35 % av artene på Rødlista, vokser i fjellet.
Arter med dårlig spredningsevne og spesielle habitatkrav er spesielt sårbare for ødeleggelse eller endringer av voksestedet. Mange moser på Rødlista tilhører denne kategorien.
Kunnskapsgrunnlaget
Den norske mosefloraen er godt undersøkt, men det finnes både geografiske områder og artsgrupper der kunnskapen om forekomsten av arter er svært mangelfull. Norge hadde en ”bryologisk gullalder” på slutten av 1800-tallet og begynnelsen av 1900-tallet med en rekke meget dyktige amatørmoseforskere, med den internasjonalt anerkjente Ingebrigt Severin Hagen i spissen. En betydelig andel av mosene i de norske universitetsherbariene ble samlet i denne perioden. Universitetsmuseenes samlinger er et viktig fundament for rødlistevurderingene. Noen av rødlisteartene er dokumentert med belegg kun fra denne perioden og mange av lokalitetene med eldre funn er ikke er oppsøkt i nyere tid.
I perioden etter den såkalte bryologiske gullalderen, var det lite innsats på kartlegging av moser, men med noen solide bidrag fra blant annet Per Størmer og Arne A. Frisvoll. Tidlig på 2000-tallet ble det gjort mange moseundersøkelser i skog i forbindelse med blant annet prosjektet Miljøregistrering i skog (MiS) (Blom mfl. 2002). Vi er nå imidlertid inne i en ny periode med relativt høy aktivitet på mosekartlegging. Vi har lagt bak oss et tiår med sterkt økende moseinteresse i Norge. I dette tiåret har flere Artsprosjekter bidratt med mye ny kunnskap om blant annet arter i fjellet, i kulturlandskapet og i nedbørrike områder på Vestlandet (Høitomt mfl. 2012, Hassel mfl. 2014, Appelgren mfl. 2016). Det har også blitt arrangert en rekke workshops under Artsprosjektparaplyen, og Norge har i tillegg fått sin egen moseklubb innordnet i Norsk botanisk forening (Høitomt og Kyrkjeeide 2015). Mange titalls personer har igjennom dette tiåret vært med på å kartlegge moser enten i regi av Artsprosjektet eller Moseklubben. Dette har ført til at mange flere personer er i stand til å finne og bestemme mange flere arter nå enn for bare ti år siden.
Vi er nå i ferd med å nærme oss et minimumsnivå av grunnleggende kunnskap om moser i de fleste habitater. Kunnskap om forekomst er fortsatt best i skog og på myr, og langt mer mangelfull i andre miljøer, men åpen kalkmark og fjell har imidlertid fått et betydelig tilfang av kunnskap de siste årene (Høitomt 2015, Høitomt og Reiso 2019).
Den solide økningen i bryologisk aktivitet kommer blant annet til uttrykk gjennom økt antall mosefunn lagt inn i Artsobservasjoner år for år. Antall poster har økt jevnt fra 10 808 poster i 2015 til 16 559 poster i 2020 (til tross for avlyste arrangementer pga. Covid-19). Antall mosearter som er kjent fra fastlandet er nå 1136, mens 1085 arter var kjent da forrige rødliste for moser ble utarbeidet (Hassel mfl. 2015). Det er en vedvarende stor interesse for moser i Norge, noe som forhåpentligvis vil resultere i bedret kunnskapsstatus for moser og enda flere som har god artskunnskap og kompetanse på moser.
I rødlisteprosessen har grunnlaget for utbredelsesdata i all hovedsak vært herbariemateriale oppbevart ved universitetsmuseene. Dette materialet er taksonomisk kvalitetssikret gjennom revisjoner gjort av Arne A. Frisvoll 1997–1998 og Kristian Hassel 2004–2005. Nyere innsamlinger er i de fleste tilfeller kontrollert av medlemmer i rødlistekomitéen for moser eller av andre eksperter i inn- og utland. Museumsmaterialet er tilgjengeliggjort via Artskart. Økningen i antall moseobservasjoner og økt bryologisk kompetanse blant frivillige kartleggere, gjør at Artsobservasjoner i økende grad tas i betraktning og særlig siden forrige rødlisting. Det foregår en løpende validering av funn innlagt i Artsobservasjoner, og bare validerte funn er lagt til grunn for rødlistevurderingene.
Kunnskapsgrunnlaget for moser er fortsatt mangelfullt når det gjelder nyere forskningsbasert viten om forekomst, habitatkrav og bestandsbiologiske forhold hos flere av moseartene på Rødlista. Det finnes lite direkte kunnskap om størrelsen på og endringer i mosepopulasjonene. Bare et fåtall arter overvåkes, og det er generelt gjort få tellinger av antall individer per lokalitet for moser. Angivelser av populasjonenes størrelse og eventuelle endringer i disse er derfor i de fleste tilfeller basert på estimater (se under).
Avgrensninger og definisjoner
Alle mosearter som er kjent fra det norske fastlandet er behandlet i rødlistearbeidet. Det ble foretatt en vurdering av 1099 arter. De resterende artene ble satt til kategori ikke vurdert NE på grunn av uavklart taksonomi og svært mangelfullt kunnskapsgrunnlag, eller til kategori ikke egnet NA fordi de er fremmede i Norge. Funn fra Svalbard er ikke inkludert i rødlistevurderingene.
I tillegg er noen få underarter vurdert. Det er kun for arter der minst én av underartene kvalifiserer til en rødlistekategori, at underarter er vurdert. For disse artene er alle underartene som forekommer i Norge tatt med, også de som er vurdert som livskraftig. Til sammen 10 underarter er vurdert. I tillegg er én underart satt til ikke vurdert NE.
De fleste moser danner tuer og matter med flere planteskudd som stammer fra samme klon. Det er derfor nødvendig å anvende såkalte sjablonger (retningslinjer) for hvordan individstørrelsen hos moser beregnes. Komitéen for den europeiske Rødlista for moser utarbeidet egne sjablonger for sitt arbeid (Bergamini mfl. 2019). For én forekomst er det tenkt et rutenett med 1 m2 ruter som dekker forekomsten, der ett individ telles som forekomsten innenfor én rute. Vi har til en viss grad fulgt Bergamini mfl. (2019), men avviker der vi mener vi har mer artsspesifikk kunnskap. Dette gjelder særlig for arter som vokser på steinblokker, trær og død ved, hvor vi har benyttet 1 dm2 rutenett for å telle individer, hvor 1 dm2 utgjør ett individ. Det samme gjelder for små kortlivete arter som vokser i til dels tette bestander på jord. I enkelte tilfeller der individene er lett å avgrense, eksempelvis ved tuevekst på stein eller trær, har vi beregnet én tue som ett individ i de videre beregningene. Disse sjablongene avviker noe fra forrige rødliste og en del arter kan derfor ha et lavere estimert individantall nå enn forrige gang.
For å beregne antall individer per lokalitet har vi benyttet kjent kunnskap om bestandsstørrelser der dette er tilgjengelig, men i de fleste tilfeller er dette ukjent. Vi har estimert antall individer per lokalitet på grunnlag av kjennskap til artens økologi og livshistorieegenskaper. Der vi har dårlig kunnskapsgrunnlag om norske forekomster, har vi brukt kunnskap fra våre naboland dersom det har vært tilgjengelig. For de fleste artene er dette vurdert individuelt. Vurderinger av antall individer per lokalitet er beskrevet nærmere i kriteriedokumentasjonen for hver enkelt art.
Rødlistekriteriene tar utgangspunkt i en vurderingsperiode som i sin tur tar utgangpunkt i artenes generasjonslengde. For de fleste moseartene mangler vi kunnskap om dette. Derfor har vi brukt anbefalingene i Bergamini mfl. (2019) som veileder: Kortlivede arter har en antatt generasjonslengde på 1-5 år - 3 generasjoner=10 år; Arter med middels levetid har en antatt generasjonslengde på 11-25 år - 3 generasjoner=50 år; Langlivede arter har en antatt generasjonslengde på 33 år - 3 generasjoner=100 år. For arter som sjelden eller aldri er funnet med sporofytter, har den lengste generasjonstiden blitt benyttet. De vanligst benyttede vurderingsperiodene for moser er 10 år, 30 år, 45 år, 60 år og 100 år.
Det har vært en del diskusjon rundt fastsettelse av minimumsverdier i forbindelse med vurderingene. Under B-kriteriet gjelder dette areal, mens under kriteriet C (og D) gjelder dette antall individer. Vi har i stor grad brukt kjent verdi (antall forekomster x antall individer per lokalitet/areal) som minimumsverdi. Dersom vi antar at alle kjente forekomster er intakte vil dette være den minste teoretiske størrelsen på utbredelsesarealet, forekomstarealet eller populasjonen. I noen tilfeller der det er helt åpenbart at artens minimumsverdier er høyere eller lavere, kan de være satt høyere eller lavere enn kjent verdi. Dette kan være nyoppdagede arter eller arter som åpenbart er underrapportert. Å sette minimum lik kjent verdi er i all hovedsak gjort for å ikke innføre ytterligere en ukjent og variabel faktor i vurderingene. Videre er det satt en maksimumsverdi. Den er satt så høyt at det er svært lite trolig at man i framtiden vil finne ut at den var for lav. Av maksimums- og minimumsverdiene regnes det ut en føre-var-vektet, såkalt beregnet verdi. I de tilfellene vi mener å ha mer eksakt kunnskap om hvilken størrelse (areal eller populasjon) som er mest sannsynlig i dette spennet, er det lagt inn en «mest trolig» verdi. Den trekker da den beregnede verdien mot den mest trolige verdien.
Norsk rødliste for naturtyper fra 2018 viser at mange naturtyper er i nokså sterk tilbakegang (f.eks. Grytnes mfl. 2018, Lyngstad mfl. 2018). Nedgang i bestandsstørrelse og areal av egnede, spesifikke habitater er estimert ut ifra tilgjengelig arealstatistikk når slik finnes, f.eks. Landsskogtakseringen. Ellers er tilgjengelig faglitteratur og egne erfaringer benyttet. For en rekke habitatspesifikke arter vil det ofte være en direkte sammenheng mellom habitatreduksjon og -forringelse og populasjonsreduksjon. Det finnes imidlertid flere eksempler på at enkeltarter er både mer eller mindre truet enn habitatet de vokser i, avhengig av hvilken del av habitatet den bruker og hvor i landet den forekommer.
Ved bruk av B-kriteriene er fragmentering et sentralt begrep. Det er imidlertid ofte vanskelig å vurdere om en mosepopulasjon er kraftig fragmentert, særlig i tilfeller der man har store hull i kunnskapen om både utbredelse og økologi. Det er i hovedsak artens utbredelse og mulighet for langdistansespredning som er benyttet i vurderingen av fragmentering. For moser har vi brukt følgende teoretiske tilnærming som veileder ved vurdering av fragmenteringsgrad:
Kraftig fragmentering: Minimumsdistanse 50 km mellom lokaliteter for arter uten kjent sporeproduksjon eller som har store sporer som ikke er tilpasset vindspredning; 100–1000 km mellom lokaliteter for arter med sporeproduksjon og vindspredning.
Trolig kraftig fragmentering: Minimumsdistanse 10 km mellom lokaliteter for arter uten sporeproduksjon eller som har store sporer som ikke er tilpasset vindspredning; 50–100 km mellom lokaliteter for arter med sporeproduksjon og vindspredning.
Klimaendringer er en mer fremtredende påvirkningsfaktor nå enn ved forrige rødlistevurdering, særlig for arter som vokser i fjellet (se under). Relativt begrenset detaljkunnskap om fjellmosenes økologi gjør at vi tok i bruk en enkel tabell for sjablongmessig vurdering (tabell 1). Denne ble brukt som veileder i mangel på bedre kunnskap og for at vurderingene innbyrdes mellom fjellartene skulle bli konsistent. Tabellene baserer seg på sammenhengen mellom habitattype og vurderingsperiode. Differensieringen i habitatenes forventede reduksjon i areal og tilstand er hentet fra Norsk rødliste for naturtyper (Grytnes mfl. 2018, Aarrestad mfl. 2018).
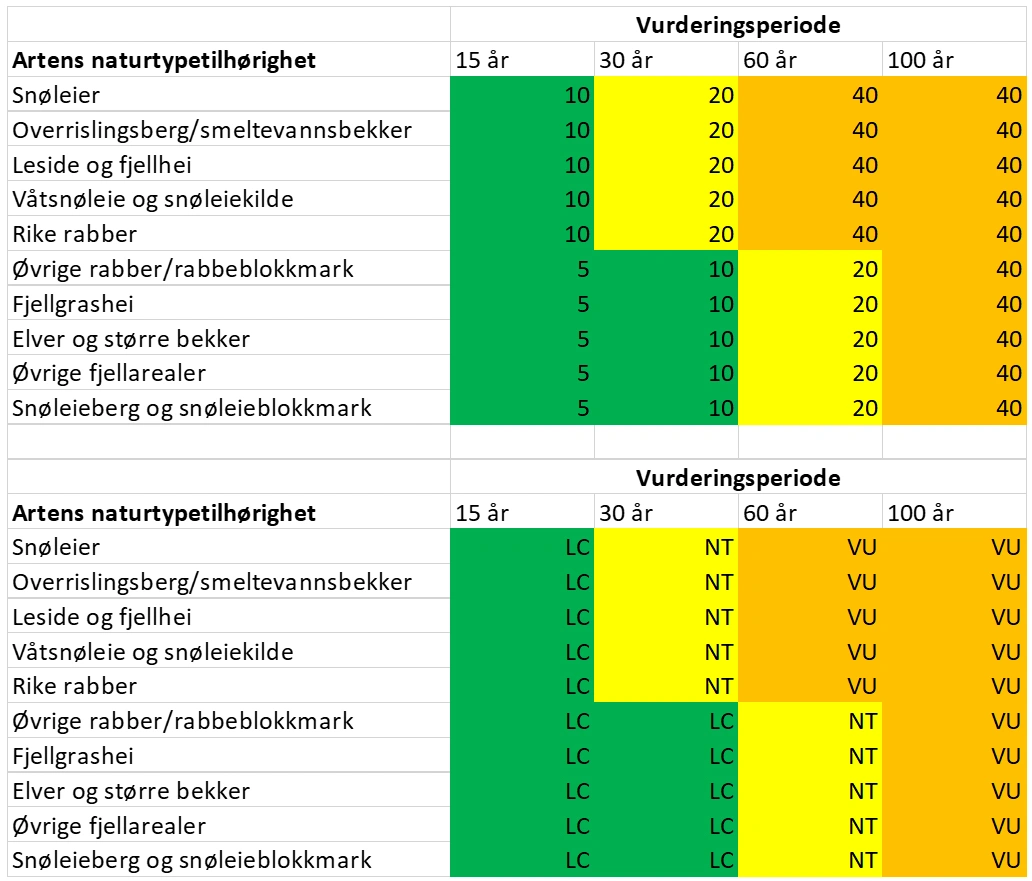
Oversikten viser sjablongmessig inndeling for å vurdere moser tilknyttet fjellhabitater. Tallene i den øverste tabellen angir forventet pågående eller framtidig prosentvis populasjonsreduksjon for ulike vurderingsperioder for arter i fjellhabitater. Nederst vises rødlistekategori tilsvarende prosentvis populasjonsreduksjon.
Antall rødlistearter
Totalt ble 1099 arter vurdert, av disse ble 777 arter vurdert til kategorien livskraftig LC, enten på grunnlag av generelle vurderinger eller på grunnlag av detaljert vurdering opp mot IUCN-kriteriene. De resterende 322 artene ble vurdert som kvalifisert til en rødlistekategori, og av disse er 243 truet (kategori kritisk truet CR, sterkt truet EN eller sårbar VU). Det er om lag hundre arter flere enn i 2015. Seks av de kritiske truete artene er vurdert som kritisk truet CR - trolig utdødd. Ingen arter er vurdert som regionalt utdødd RE. Nitten arter er vurdert i kategorien datamangel (DD).
Av de 10 underartene som er vurdert er sju rødlistet og tre vurdert som livskraftig LC.
Årsak til rødlisting
De fleste moseartene er rødlistet med grunnlag i C-kriteriet liten populasjonsstørrelse og pågående nedgang (41 %) og D-kriteriet svært liten populasjonsstørrelse (23 %). B-kriteriet begrenset utbredelsesområde og/eller forekomstareal (18 %) og A-kriteriet reduksjon i populasjonsstørrelse (16 %) er brukt i liten grad. Bruken av A-kriteriet, og da i første rekke A3-kriteriet, har økt siden revisjonen i 2015, noe som skyldes en mer helthetlig tilnærming til klimaendringene og hvordan de er forventet å påvirke fjellnaturen vår i tiårene som kommer. For mange arter har vi estimert reduksjon i arealet med tilgjengelig habitat og nedgang i populasjonsstørrelsen. I slike tilfeller er oftest C-kriteriet utslagsgivende fordi det gir en høyere truethetskategori enn B-kriteriet. For sjeldne arter der vi har estimert svak eller ingen nedgang, er de vurdert etter D1-kriteriet, hvor populasjonsstørrelsen er avgjørende for hvilken kategori arten plasseres i. Mange mosearter med få forekomster og spesialisert økologi er vurdert etter D-kriteriet.
Påvirkning på habitat, som vil si arealbruksendring og endret økologisk tilstand i naturtyper som følge av menneskelige inngrep og aktivitet, er den aller viktigste påvirkningsfaktoren for moser. Deretter utgjør klimatiske endringer en betydelig trussel, og forurensing er den tredje viktigste påvirkningsfaktoren. Disse tre omtales her, mens detaljer for hver enkelt art er angitt i kriteriedokumentasjonen for arten.
Av påvirkning på habitat er det endringer i landbruket som er den viktigste påvirkningsfaktoren. Opphør av beite utgjør den største av underkategoriene innen landbruk. Opphør av bruk, som slått og beite, fører til at semi-naturlig jordbruksmark gror igjen med busker og trær. Dette påvirker mosefloraen negativt. Dette gjelder blant annet sjeldne pionérmoser med sørvestlig utbredelse knyttet til kystlynghei, f.eks. Campylopus brevipilus og Leptodontium flexifolium (Blockeel mfl. 2014). Semi-naturlige-, boreonemorale tørrenger, som inneholder mange sjeldne, sørøstlige mosearter, gror nå raskt igjen som følge av opphør av beiting (Grytnes mfl. 2018, Reiso og Haugan 2010). Det finnes lite forskning som dokumenterer hastigheten på denne gjengroingen, men det er observert at bestander av flere sjeldne moser er går tilbake som følge av skyggeeffekter fra busk- og krattvegetasjon, ofte med en kombinasjon av stedegne og fremmede arter.
En annen viktig påvirkningsfaktor innen underkategori landbruk, er skogbruk, med flatehogst, åpne hogstformer og treslagsskifte/plantasjer som de viktigste påvirkningsfaktorene. Mange habitatspesialiserte moser har sin utbredelse knyttet til bergvegger, kløfter, bekkedaler og sumper i og omkring naturskog. Det moderne skogbruket har, særlig på grunn av store flatehogster, påvirket mosefloraen negativt både direkte og indirekte. Direkte påvirkning har trolig skjedd først og fremst på grunn av endret mikroklima i områder med flatehogst. Moderne skogbruk fører til mindre fuktig og mer ustabilt mikroklima i perioden etter hogsten (Framstad mfl. 1995), blant annet ved at solinnstrålingen på bergvegger og i sumpmiljøer øker. De siste par tiårene har imidlertid stående volum av trær i alle aldersklasser økt, i granskog, furuskog og lauvskog. Også mengden av død ved har økt (Larsson og Hylen 2007, Storaunet og Rolstad 2015). Denne økningen er imidlertid ikke jevnt fordelt i skoglandskapet, og siden arealandelen naturskog blir stadig lavere, vil mye av den nye dødveden trolig bli dannet som følge av sammenbrudd i nokså ensaldrete skogbestand. Det er usikkert om denne dødveden er av egnet kvalitet for mange truete moser, og om moser med dårlig spredningsevne i det hele tatt rekker å spre og etablere seg i løpet av tidsvinduet substratet er tilgjengelig. Mange råtevedmoser (særlig levermoser) er avhengige av slike livsmedier for å gjennomføre sin livssyklus. Effekter av fragmentering av naturskog på mosefloraen er dårlig undersøkt. Mange av artene som er knyttet til råteved har imidlertid hovedsakelig ukjønnet formering ved grokorn. Slike arter har dårlig spredningsevne og viser ofte en klumpet utbredelse på landskapsnivå (Korpelainen mfl. 2011, Laaka-Lindberg mfl. 2006).
Drenering og grøfting av myr, sumpskog og annen våtmark til landbruks- og skogbruksformål har tidligere hatt et betydelig omfang. Dette har først og fremst vært konsentrert til lavlandsområdene og 25 % av det opprinnelige myrarealet under skoggrensa i Norge er drenert (Moen 1995). Dette har utvilsomt ført til en reduksjon i forekomstareal og mer fragmentert forekomst for mange sjeldne myrmoser, inkludert rødlistede arter. Nylig ble nydyrking av myr forbudt (Landbruks- og matdepartementet 2020). Allikevel vil tidligere drenerte habitater fortsette å endre seg over tid som følge av såkalt endringsgjeld og arter kan trues lenge etter inngrepet ble utført (Kuussaari mfl. 2009).
Habitatpåvirkning i limniske miljø utgjør en viktig påvirkningsfaktor for mange moser. Det er spesielt oppdemming og vannstandsregulering som er den antatt viktigste påvirkningsfaktoren. Slike inngrep har negativ påvirkning på moseforekomster gjennom direkte habitatødeleggelse, men påvirker også moser gjennom endrete fuktighetsforhold i overgangen mellom vann og land. Kunnskapsnivået om effekter av vassdragsregulering på moser er begrenset; særlig gjelder dette for sjeldne og truete arter. Det er gjort en liten undersøkelse av fakkeltvebladmose (Hassel 2018). Utbygging av mini- og småkraftverk i bratt terreng kan ha betydelig effekt på moser som er avhengig av stabil, høg, fuktighet (Hassel og Holien 2007, 2008). Særlig knytter det seg stor usikkerhet til i hvilken grad bergveggsmoser i kløfter og gjel påvirkes negativt av tørrlegging eller redusert vannføring. En vesentlig grunn til dette er en ond sirkel hvor kunnskapen om mosefloraen i disse habitatene er mangelfull og kartleggere ikke kan artene, som fører til at rødlistede moser bare i svært liten grad fanges opp i forbindelse med konsekvensutredninger for småkraftutbygging (Gaarder og Høitomt 2014).
Andre årsaker til arealbruksendring er utbyggingspress med særlig boligbebyggelse inkludert fritidsboliger, og infrastruktur. Dette har trolig betydelig negativ effekt på mosepopulasjoner, både ved direkte habitatødeleggelse og ved fragmentering av habitater. Dette gjelder spesielt for habitatspesifikke og varmekjære moser med sørlig utbredelse. Effekten på mosefloraen antas å være størst i lavlandet i Sør-Norge, i områder med høy befolkningstetthet. Dokumentasjonen av dette er imidlertid mangelfull. Mange arter som er knyttet til semi-naturlige, boreonemorale tørrenger (se over) finnes også i åpen grunnlendt kalkmark, som er naturlig åpne arealer på grunt jordsmonn og som gjerne forekommer som en smal stripe mellom strandberg og skogsmarka innenfor, eller på knauser og nær skrenter. Arealbrukspresset på kystnære områder er årsak til tilbakegang for disse moseartene også i denne naturtypen.
Klimaendringer er årsak til at flere arter er rødlistet nå enn forrige gang, noe som skyldes ny kunnskap og en mer helhetlig tilnærming til denne påvirkningsfaktoren enn tidligere. Utgangspunktet er at en forventet oppvarming på 2,5 grader de neste 50 år gir gunstigere forhold for etablering av skog på opptil 85 % av arealet som i dag er snaufjell (Grytnes mfl. 2018). Det er en forsinkelse i disse endringene, men følgene vil allikevel bli alvorlige. Følger av storskala klimaendringer for utbredelsen av mosearter i Norge ble til en viss grad vurdert i 2015 og effektene av temperaturøkning og økning i frekvensen av ekstremvær i form av lengre perioder med tørke vurdert som negativ for noen av våre fjellarter. Konsekvensene klimaendringene vil ha på moser er nå vurdert for alle fjellarter og arter med store deler av sin utbredelse over tregrensa. Det er ventet at det vil bli betydelig høyere temperaturer, kortere snødekkevarighet og generelt noe fuktigere vær, særlig i Nord-Norge (Norsk klimaservicesenter 2021). Dette vil gi endringer i vekstsesongen og dermed påvirke arter som er tilpasset kort vekstsesong og kaldt klima (Aarrestad og Grytnes 2018). Gjengroing og høyere karplantevegetasjon kan ventes, noe som på sikt vil utkonkurrere moser. En slik effekt er allerede dokumentert fra Sverige (Alatalo mfl. 2019). Videre vil permafrost og permanente snøfonner smelte raskere (Kjøllmoen mfl. 2010, Koch mfl. 2014, Nesje mfl. 2012, Wolden 2015) og jevn tilførsel av vannsig i høyfjellet vil med tiden avta, blant annet forventes snøleier å bli kraftig påvirket (Aarrestad mfl. 2018). Tørrere forhold vil ha negative konsekvenser for snøleiearter og andre moser som er tilpasset jevn fuktighet (Björk og Molau 2007, Sandvik og Odland 2014). Moser i fjellet antas å bli påvirket negativt både gjennom direkte mortalitet som følge av økte temperaturer og konkurranse fra mer varmekrevende arter. For flere arter vil trolig begge disse faktorene virke sammen. I rødlistesammenheng blir utslagene mest alvorlig for langlivete arter i smeltevannsmiljøer, der veldig mange arter nå blir vurdert som sårbare VU.
Det er også sannsynlig at klimaendringene vil bedre forholdene for mange mosearter i Norge. Arter med sørlig utbredelse vil etter hvert kunne få økte populasjoner. Disse endringene er mye vanskeligere å forutsi enn endringene i fjellet, fordi naturtypene ikke nødvendigvis endres like drastisk som når vegetasjonssonene flytter seg oppover i fjellet. Det kreves trolig overvåkningsopplegg for aktuelle arter for å finne ut om en forventet ekspansjon finner sted og hvor raskt det eventuelt går. Dette er uansett problemstillinger som må vurderes mer grundig ved kommende rødlisterevisjoner.
Forurensing er også en aktuell påvirkningsfaktor for noen mosearter. Flere sjeldne moser er barkboende (epifytter) på naturlig forekommende eller innplantete varmekjære løvtrær i Sør-Norge. Trolig har flere av disse artene tidligere vært i tilbakegang i Sør-Norge som følge av sur nedbør, men dokumentasjonen på dette er mangelfull. De siste tre tiårene har imidlertid forsuringen gradvis nærmest opphørt i fleste steder, i hvert fall i sentrale og vestlige deler av Europa. Flere steder i Europa er det samtidig dokumentert at epifyttfloraen ekspanderer i tidligere forsurete områder. Dette har også ført til at flere arter også inntar nye områder (Blockeel mfl. 2014). Mange steder, inkludert i Norge, kan en slik fremrykning forsterkes av det økende løvtreoppslaget i mange områder. I likhet med i Sør-Sverige (Gunnarsson mfl. 2000) er det rimelig å anta at forsuringen av torv og vann i rikmyrer i Sør-Norge også har resultert i tilbakegang for flere myrmoser som krever mineralnæringsrike vekstforhold. Forsuringen av elver og vassdrag i Sør-Norge har trolig også hatt en negativ påvirkning på utbredelse og forekomst av flere mineralnæringskrevende vannmoser.
Endringer fra 2015 til 2021
Det har skjedd store endringer på Rødlista 2021 sammenlignet med 2015. De største endringene er knyttet til at mange fjellarter nå har blitt vurdert med pågående og framtidige klimaendringer som påvirkningsfaktor. Dette betyr at om lag 60 nye arter med sin hovedutbredelse i fjellet nå er rødlistet. Noen arter fra andre naturtyper er også nye på Rødlista og i tillegg har noen nyoppdagete arter tilkommet. Til sammen teller Rødlista for moser i 2021 hele 85 flere arter enn i 2015. Til sammen er nå 322 norske moser rødlistet.
I alt har 85 arter som var rødlistet i 2015 endret rødlistekategori til Rødlista i 2021. Totalt er 19 arter vurdert til lavere kategori eller tatt ut av rødlista. Av disse er 10 vurdert til kategori livskraftig LC nå i 2021. De resterende 66 artene er plassert i en høyere rødlistekategori enn i 2015. I tillegg har 69 arter som var vurdert som livskraftige LC i 2015, nå blitt rødlistet. Antall arter i kategorien DD – datamangel er redusert fra 44 i 2015 til 19 i 2021.
Ny kunnskap om arters utbredelse og påvirkningsfaktorer en den viktigste enkeltårsaken til endring i rødlistekategori fra 2015 til 2021. Ny kunnskap har tilkommet gjennom en økt feltinnsats, både fra profesjonelle og fra amatørbryologer. I mange tilfeller har kunnskapen økt både om enkeltarter og om utviklingen for deres habitater. Større sikkerhet i vurderingene av bestandsstørrelser har i flere tilfeller ført til en mer alvorlig truethetskategori, men i enkelte tilfeller har ny kunnskap ført til nedjustering av truethetskategorien. I tillegg kommer den store andelen med fjellarter som mer eller mindre sjablongmessig nå er vurdert. At dette grepet blir gjort akkurat nå skyldes både at det var ressurser tilgjengelig til en slik gjennomgang og at vi vurderer dokumentasjonen av sannsynlige effekter av klimaendringer i framtid som gode nok til å kunne benytte kriteriet A3 for disse artene. Fjellarter med begrenset utbredelse eller små populasjoner vil som i 2015 ha B-, C- og/eller D-kriteriet som gjeldende kriterium.
Den føre-var-justerte beregnede verdien som ble innført i årets vurderingsarbeid har ført til noen få kategoriendringer. Det samme har endringer i sjablongene for individstørrelse. Ny kunnskap om enkelte av artene har også ført til kategoriendringer, men det er viktig å presisere at det stort sett er den nye kunnskapen i seg selv og ikke mistanke om faktiske populasjonsendringer som fører til disse justeringene. Ny kunnskap om hvordan fjellene våre vil påvirkes av klimaendringer er den viktigste årsaken til at mange nokså tallrike og vidt utbredte fjellarter nå har kommet inn på rødlista. Ny eller endret kunnskap er oppgitt som viktigste årsak til endring i kategori for arter som var på Rødlista i 2015 og som fortsatt er der i 2021.
Ekspertkomiteen
Ekspertkomiteen har bestått av Torbjørn Høitomt (leder), Hans H. Blom, Kristian Hassel, John Gunnar Brynjulvsrud og Magni Olsen Kyrkjeeide. Artene ble fordelt mellom ekspertene basert på erfaring og kompetanse på artene, med Høitomt og Blom som ansvarlige for flest arter. Kyrkjeeide og Høitomt har kvalitetssikret arbeidet.
Takk
Takk til Anders Breili, Jon Inge Johnsen og Tom Hellik Hofton for nyttige innspill og diskusjoner underveis i prosessen. Takk også til rødlisteteamet i Artsdatabanken for godt samarbeid.
Referanser
Alatalo J, Jägerbrand A, Chen S, Sun S-Q og Molau U (2019). Bryophyte cover and richness decline after 18 years of experimental warming in Alpine Sweden. EcoEvoRxiv.org 2019
Appelgren L, Carlsson P og Homble K (2016). Scopelophila ligulata recorded for the first time in northern Europe. Lindbergia 39: 12–19, 2016.
Aarrestad PA og Grytnes J-A (2018). Fjellhei, leside og tundra, Fjell og berg. Norsk rødliste for naturtyper 2018. Artsdatabanken, Trondheim. Hentet 12.04.2021 fra: https://artsdatabanken.no/RLN2018/43
Aarrestad PA, Evju M, Høitomt T, Ihlen P og Grytnes J-A (2018). Snøleie, Fjell og berg. Norsk rødliste for naturtyper 2018. Artsdatabanken, Trondheim. Hentet 12.04.2021 fra: https://artsdatabanken.no/RLN2018/46
Bergamini A, Bisang I, Hodgetts N, Lockhart N og van Jacques R (2019). Recommendations for the use of critical terms when applying IUCN red-listing criteria to bryophytes. Lindbergia. 42: linbg.01117
Björk RG og Molau U (2007). Ecology of Alpine Snowbeds and the Impact of Global Change. Arctic, Antarctic, and Alpine Research 39: 34-43
Blockeel TL, Bosanquet SDS, Hill MO og Preston CD (2014). Atlas of British & Irish Bryophytes. The distribution and habitat of mosses and liverworts in Britain and Ireland. Pisces Publications, Newbury
Blom HH, Hassel K og Prestø T (2002). Moser. I: Gjerde I, Baumann C (red.) Miljøregistreringer i Skog - biologisk mangfold. Norsk institutt for skogforskning, s. 52-60
Framstad E, Bendiksen E, Flatberg K I, Frisvoll AA, Holien H, Høiland K, Prestø T og Svalastog D (1995). Planter i boreal skog: effekter av lokale økologiske faktorer, skogsdrift og omgivelser på artsmangfoldet. Aktuelt fra Skogforsk 32
Gaarder G og Høitomt T (2014). Etterundersøkelse av flora og naturtyper i elver med planlagt småkraftutbygging. Foreløpige vurderinger. Miljøfaglig Utredning notat 2014-20
Grytnes J-A, Evju M, Høitomt T, Ihlen PG og Aarrestad PA (2018). Fjell og berg. Norsk rødliste for naturtyper 2018. Artsdatabanken. Hentet 12.04.2021 https://www.artsdatabanken.no/Pages/258605
Gunnarsson U, Rydin H og Sjörs H (2000). Diversity and pH changes after 50 years on the boreal mire Skattlosbergs Stormosse, Central Sweden. Jornal of Vegetation Science 11: 277-286
Hassel K, Appelgren L, Blom HH, Flynn KM, Gaarder G, Heegaard E, Høitomt T, Jordal JB, Lima M, Söderström L og Wangen K (2014). Colura calyptrifolia a new oceanic liverwort to Norway and Scandinavia. Lindbergia 37: 1-5
Hassel K og Holien H (2007). Biologisk kartlegging av fossesprutsoner i kommunene Høylandet, Stjørdal og Verdal i Nord-Trøndelag. NTNU, Vitenskapsmuseet Rapport botanisk serie, 1-28
Hassel K og Holien H (2008). Biologisk kartlegging av fossesprutsoner i kommunene Namsos, Namdalseid og Steinkjer i Nord-Trøndelag. NTNU, Vitenskapsmuseet Rapport botanisk serie, 1-35
Hassel K, Prestø T og Schmidt NM (2012). Bryophyte diversity in high and low arctic Greenland. Establishment of permanent monitoring transects and bryophyte mapping in Zackenberg and Kobbefjord 2009-2010. Scientific Report from DCE - Danish Centre for Environment and Energy, s 1-46
Hassel K, Blom HH, Høitomt T og Halvorsen R (2015). Moser (Anthocerotophyta, Marchantiophyta, Bryophyta). Norsk rødliste for arter 2015. Artsdatabanken <http://www.artsdatabanken.no/Rodliste/Artsgruppene/Moser>. Nedlastet 12.04.2021
Hassel K (2018). Hva er effekten av småkraftverkutbygging på moser som vokser på liggende dødved? En populasjonsstudie av fakkeltvebladmose Scapania apiculata. – Blyttia 76: 127-133.
Høitomt T, Appelgren L, Lönnell N, Lye K og Hassel K (2012). Pyramidula tetragona (Brid.) Brid. rediscovered in Fennoscandia and new to Norway. Lindbergia 35: 33-39
Høitomt T (2015). Kartlegging av moser på åpen kalkmark i Oslo og Akershus. BioFokus-rapport 2015-18. 27 s.
Høitomt T og Kyrkjeeide M (2015). En norsk moseklubb blir til. Blyttia. 73: 48-50.
Høitomt T og Reiso S (2019). Kartlegging av åpen kalkmark i Buskerud 2018. BioFokus-rapport 2019-7. Stiftelsen BioFokus. Oslo
Kjøllmoen B, Andreassen LM, Elvehøy H, Jackson M og Giesen RH (2010). Glaciological investigations in Norway in 2009. NVE Report 2 2010, 85 s.
Koch J, Clague JJ og Osborn G (2014). Alpine glaciers and permanent ice and snow patches in western Canada approach their smallest sizes since the mid-Holocene, consistent with global trends. The Holocene 24: 1639-1648
Korpelainen H, von Cräutlein M, Laaka-Lindberg S og Huttunen S (2011). Fine-scale spatial genetic structure of a liverwort (Barbilophozia attenuata) within a network of ant trails. Evolutionary Ecology 25: 45-57
Kuussaari M, Bommarco R, Heikkinen RK, Helm A, Krauss J, Lindborg R, Öckinger E, Pärtel M, Pino J, Rodà F, Stefanescu C, Teder T, Zobel M og Steffan-Dewenter I (2009). Extinction debt: a challenge for biodiversity conservation. Trends in Ecology & Evolution 24: 564- 571.
Laaka-Lindberg S, Korpelainen H, Pohjamo M (2006). Spatial distribution of epixylic hepatics in relation to substrate in a boreal old-growth forest. Journal Hattori Botanical Laboratory 100: 311-323
Landbruks- og matdepartementet (2020). Forskrift om nydyrking. https://lovdata.no/dokument/SF/forskrift/1997-05-02-423. Besøkt 01.08.2020.
Larsson JY og Hylen G (2007). Skogen i Norge. Statistikk over skogressurser i Norge registrert i perioden 200-2004. Viten fra Skog og Landskap
Lindgaard A og Henriksen S (2011). Norsk rødliste for naturtyper 2011. Artsdatabanken, Trondheim
Longton RE (1988). The biology of polar bryophytes and lichens. Cambridge University Press, Cambridge
Lyngstad A, Brandrud TE, Moen A og Øien DI (2018). Norsk rødliste for naturtyper 2018 – Våtmark. – NTNU Vitenskapsmuseet naturhistorisk notat 2018-15: 1-117.
Moen A (1995). The Norwegian national plan for mire reserves: methods, criteria and results. Gunneria 70: 156-176
Nesje A, Pilø LH, Finstad E, Solli B, Wangen V, Ødegård RS, Isaksen K, Støren EN, Bakke DI og Andreassen LM (2012). The climatic significance of artefacts related to prehistoric reindeer hunting exposed at melting ice patches in southern Norway. The Holocene 22: 485-496
Norsk klimaservicesenter. 2021. Klimaframskrivninger. Nedlastet: 5.4.2021. https://klimaservicesenter.no/climateprojections?index=air_temperature&period=Annual&scenario=RCP85&area=NO
Puttick MN, Morris JL, Williams TA, Cymon JC, Edwards D, Kenrick P, Pressel S, Wellman CH, Schneider H, Pisani D og Donoghue PCJ (2018). The interrelationships of land plant and the nature of the ancestral embryophyte. Current Biology. 28: 733-745
Reiso S og Haugan R (2010). Kartlegging av rødlistede kalklav i utvalgte verneområder i Bamble og Porsgrunn kommuner. Vurdering av trusler og skjøtselsbehov. BioFokus-rapport 2010-14
Reiso S og Høitomt T (2015). Kartlegging av åpen kalkmark i Hole og Ringerike kommuner, Buskerud 2014. BioFokus-rapport 2015-6
Reiso S, Høitomt T og Thylen A (2014). Kartlegging av åpen kalkmark i Buskerud, Vestfold, Telemark, Oppland og Hedmark 2013. BioFokus-rapport 2014-8
Reiso S, Abel K, Hofton TH, Høitomt T og Olberg S (2011). Åpen grunnlendt kalkmark i Oslofeltet. Innspill til faggrunnlag for handlingsplan. BioFokus-rapport 2011-44
Sandvik SM og Odland A (2014). Changes in alpine snowbed-wetland vegetation over three decades in northern Norway. Nordic Journal of Botany 32: 377-384
Storaunet KO og Rolstad J (2015). Mengde og utvikling av død ved i produktiv skog i Norge. Med basis i data fra Landsskogtakseringens 7. (1994-1998) og 10. takst (2010-2013). Skog og landskap, Oppdragsrapport 06/2015
Wolden G (2015). Urgamle snøfonner smelter i rekordfart. Gemini. http://gemini.no/2015/01/urgamle-snofonner-smelter-i-rekordfart/
Siden siteres som:
Høitomt T, Blom HH, Kyrkjeeide MO, Hassel K, Brynjulvsrud JG (2021). Artsgruppeomtale moser (Anthocerotophyta, Marchantiophyta og Bryophyta). Norsk rødliste for arter 2021. Artsdatabanken. https://www.artsdatabanken.no/rodlisteforarter2021/Artsgruppene/Moser Nedlastet <dag/måned/år>